|
aus:
Beiträge
zur Heimatkunde der Stadt Schwelm und ihrer Umgebung, Neue Folge, 52
(2003): 7-27, Abb. 1-16, Tab. 1; Schwelm.
(verändert)
Geologisch-paläontologische
Untersuchungen am Zuckerberg in Ennepetal (Givetium, nordwestliches
Sauerland)
Lutz Koch und Ulrich Lemke
Einleitung
Östlich von
Schwelm, auf Ennepetaler Stadtgebiet zwischen
Büttenberg und Oelkinghausen, stehen am Nordhang des Zuckerberges (290
m NN) in einem alten fast völlig verfallenen bzw. überwachsenen
Steinbruch mitteldevonische Sedimente der Honsel-Formation an, die
stellenweise fossilreich sind. Vom Arbeitskreis Kluterthöhle
durchgeführte Grabungen auf der Suche nach vermuteten Hohlräumen im
Kalkstein der Schichtfolge lieferten frisches Gesteinsmaterial, das auf
seine Fossilführung untersucht wurde. Darüber hinaus wurden auch die
anstehenden Gesteine im Liegenden (bis zur Eisenbahnstrecke) und im
Hangenden (Waldgebiet oberhalb des Aufschlusses) beprobt, so dass ein
geologisches Gesamtprofil des Zuckerberges erstellt werden konnte. Die
Erkenntnisse werden ergänzt durch Ergebnisse niedergebrachter Bohrungen
im Rahmen der Planung der Bundesstraße 7e, die parallel zur
Eisenbahnstrecke längs des Zuckerberges verlaufen sollte1.
|

|
|

|
|
Alter Steinbruch am
Nordhang des Zuckerberges.
|
Die Schichtfolge
Auf der Geologischen Karte von Nordrhein-Westfalen 1: 25 000 (Blatt
4709 Barmen) wurde die Fundstelle von Paeckelmann in Fuchs &
Paeckelmann (1979) als zu den „Oberen Honseler Schichten"
gehörend kartiert sowie der ehemalige Steinbruch als Fossilfundpunkt2
ausgewiesen. Aufgrund des häufigen Vorkommens der Trilobiten-Art Dechenella
burmeisteri, des Nachweises der Brachiopoden Undispirifer
sp. und Crurithyris inflata sowie des Ostracoden Bairdiocypris
sp. kann diese Einstufung vorbehaltlich bestätigt werden. Das
Auffinden von Spinocyrtia (Carpinaria) ascendens
spricht für Honsel-Formation in ihrer Gesamtheit. Andererseits enthält
aber die Schichtfolge auch Formen, die typisch sind für Unteres
Mitteldevon (Eifelium): z. B. Cyrtina heteroclita, Helaspis
plexa und Dicamara cf. plebeja. Einige weitere zur
Altersbestimmung geeignete Brachiopoden wie Atrypiden3 sind
nicht ausreichend gut erhalten, um sie eindeutig auf Art- bzw.
Unterart-Ebene bestimmen zu können. Schließlich blieben durchgeführte
Conodonten4-Untersuchungen ohne entsprechende Nachweise.
Der Kalksteinhorizont am Zuckerberg ist Bestandteil der Ennepetaler
Plateau-Riffe, die aus zwei ca. 10-12 Meter mächtigen Kalklinsen
bestehen, unterbrochen von tonig-sandigen Sedimenten. Diese
Kalksteinlagen finden sich am Klutertberg (Koch 1992), im Heilenbecker
Tal an der Wassermaus, auf der Hardt, am Timpen, an der Kahlenbecke, im
Hembecker Tal sowie am Zuckerberg. Auch bei dem 1500 Meter südwestlich
des Zuckerberges gelegenen Eisenbahneinschnitt bei Martfeld handelt es sich um Gesteine der Oberhonsel-Formation, die hier
direkten Kontakt zum Schwelmer Massenkalk im Hangenden besitzen.
Die von May (1986: 33) neu definierte Honsel-Formation (vgl. auch
Weddige 1996: 281) besteht überwiegend aus grauen Feinsandsteinen. In
die sandsteinreiche Schichtenfolge sind siltig-tonige Lagen sowie
Korallenkalke eingeschaltet. Während die Unterhonsel- Formation zudem
noch Rotschiefer-Horizonte enthält, ist die Oberhonsel-Formation frei
von Rotschiefer-Einlagerungen.
Über die Schwierigkeit, lokal begrenzte Korallenkalk-Horizonte
stratigraphisch miteinander zu vergleichen, berichtet May (1986: 43). So
kann auch nichts darüber ausgesagt werden, ob die Ennepetaler
Kalklinsen z. B. mit den Kalk-Horizonten im Blattbereich Iserlohn
parallelisiert werden können.
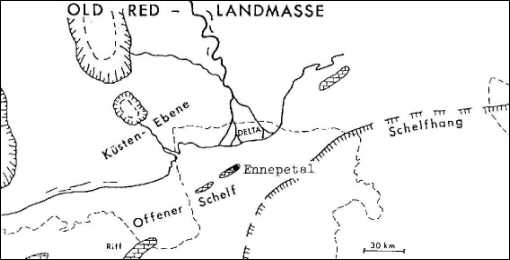
Entstanden ist die Schichtenfolge vor ca. 380 Millionen Jahren in
einem flachen Meer auf dem inneren Schelf des Old-Red-Kontinentes in
bewegtem Wasser bei einer Tiefe von nur wenigen Metern. Die
Sandschüttungen kamen aus einem nördlich gelegenen Delta, von wo aus
auch zahlreiche Pflanzenreste in den Schelf befördert wurden.
|
|
|
Abb. 1: Paläogeographische Rekonstruktion
des Rheinischen Schelfs zur Zeit des Mitteldevon.
Die Küstenlinie liegt bei etwa 10° bis 20° südlicher Paläobreite
(aus Koch 1992, nach Langenstrassen
1982). |
Die am Zuckerberg aufgeschlossenen Sedimente enthalten vom Liegenden
(Eisenbahnstrecke) zum Hangenden (Wald oberhalb des Steinbruchs)
folgende Schichtglieder:
[280 m NN »Wald«]
Obere Siltsteinfolge: 10 m sehr harter
ungeschichteter graubrauner grober Sandstein mit eingelagerten braunen
mulmig verwitternden Siltstein-Lagen mit z. T. lagenweise
angereicherten Fossilien.
Tonkalkstein: 0,2 m harter blaugrauer sandig-toniger Kalkstein
(ohne Riffbildner), teilweise Kalkknollen.
Obere Sandsteinfolge: 4 m feinkörniger grau-blauer bis
grau-grüner bankig-plattiger Feinsandstein, der mehr oder weniger
kalkhaltig sein kann und gelegentlich gut konservierte Fossilien
enthält.
Obere Kalksteinfolge:
8 m dunkler, harter blauschwarzer
fossilreicher Kalkstein mit Rifffauna.
Untere Sandsteinsteinfolge:
20 m feinkörniger grauer bis
brauner bankig-plattiger Feinsandstein, teilweise mit laminierter
Schichtung, nach oben zunehmend kalkhaltig.
Untere Siltsteinfolge: 20 m grober Sandstein und graubrauner
sandiger Tonstein sowie brauner mulmig verwitternder Siltstein mit z.
T. lagenweise angereicherten Fossilien.
[220 m NN »Eisenbahnstrecke«]
Bei einer in einer Höhe von 229 m NN durchgeführten Bohrung5
unterhalb des Steinbruchs setzt sich die Siltsteinfolge fort und geht
bei – 10,40 m in eine Wechsellagerung von Sandstein und unreinem
Kalkstein über. Sie erreicht bei einer Teufe von – 15,30 m die Untere
Kalksteinfolge (Korallenkalk).
[229 m NN »Bohrung«]
± 0 m
-- Humus.
– 0,25 m -- Untere
Siltsteinfolge: Ton, Schluff und
stark verwitterter
Sandstein.
–10,40 m -- sandig-toniger Kalkstein.
–10,80 m -- Wechsellagerung von Kalk- und Sandstein.
–15,30 m -- Untere Kalksteinfolge: Korallenkalk.
Diese Abfolge mit den beschriebenen Sedimenttypen spiegelt die
unterschiedlichen Faziesbereiche während der Honsel-Zeit wider, die von
Çinar (1978) auch für die Honsel-Formation in der Lüdenscheider Mulde
festgestellt wurden. Demnach sorgt eine lang anhaltende
sedimentologische und paläogeographische Entwicklung mit rhythmisch
wechselnden Ablagerungsbedingungen für das Entstehen und Absterben der
Kleinriffe: Zunächst wird das tonig-siltige Substrat besonders von
Bryozoen und Brachiopoden besiedelt. Danach folgen sich flach
ausbreitende Bödenkorallen. Damit ist die Grundlage für die Siedlung
von Riffbildnern (Korallen und Stromatoporen) geschaffen.
Die geringe Mächtigkeit der Ennepetaler Kalksteinhorizonte und die
sandig-tonigen Zwischenlagen zeigen, dass das Riffwachstum immer wieder
durch Schüttungen von Norden gestört und nach einem relativ kurzen
Zeitraum wieder beendet wurde. Am Zuckerberg-Steinbruch ist die obere
Kalklinse aufgeschlossen, und selbst die unterschiedlichen Sedimente im
Hangenden, die das einstige Riff erstickten, zeigen einen häufigen
Fazieswechsel mit sich rasch ändernden Lebensbedingungen.
|
Mächtigkeit |
Gestein |
Fossilführung |
|
ca. 10 Meter |
harter ungeschichteter graubrauner grober Sandstein mit
eingelagerten braunen mulmig verwitternden Siltsteinen
|
Fauna z. T. aus Lesesteinen
Pflanzen: unbestimmbare Pflanzenreste
Ichnofossilien: Chondrites sp.
Crinoiden (Seelilien): Stielglieder
Rugose Korallen: Heliophyllum sp.
Tabulate Korallen (Bödenkorallen): Favosites sp., Heliolites
sp.
Bryozoen (Moostierchen): Fenestella sp.
Brachiopoden (Armfüßer): Spinatrypa aspera (Schlotheim
1813), Spinocyrtia (Carpinaria) ascendens (Spriestersbach
1935), Cyrtina heteroclita (Defrance 1828), Undispirifer
sp., ?Subcuspidella sp., Athyris sp., Dicamara
cf. plebeja (Sowerby in Sedgwick & Murchison 1840), Helaspis
plexa (Wolfart 1956)
Gastropoden (Schnecken): Straparollus sp., Loxonema
sp., Platyceras sp.
Muscheln: Ptychopteria (Actinopteria) reticulata
(Goldfuss), Leptodesma wupperana Herm. Schmidt 1909
Hyolithen: "Theca" sp.
Cephalopoden (Kopffüßer): "Orthoceras"
sp., Rutoceras sp.
Ostracoden (Muschelkrebse): indet.
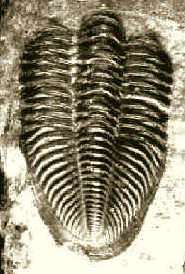
Trilobiten: Dechenella burmeisteri Rud. Richter 1909
|
|
ca. 0,2 Meter |
blaugrauer sandiger Kalkstein |
Brachiopoden (Armfüßer): Crurithyris
inflata (Schnur 1853)
Tentaculiten: indet.
Trilobiten: Dechenella sp.
nicht näher bestimmbare Kleinfauna
|
|
ca. 4 Meter |
feinkörniger grau-blauer bis grau-grüner bankig-plattiger
Feinsandstein, teilweise kalkhaltig |
Pflanzen: Asteroxylon
elberfeldense Kräusel & Weyland 1926, Aneurophyton
germanicum Kräusel & Weyland 1923, Protopteridium
thomsonii Kräusel & Weyland 1932
Ichnofossilien: Chondrites sp., Grabspuren indet.,
Fraßspuren in Holzresten
Anneliden (Ringelwürmer): Spirorbis sp.
Bryozoen (Moostierchen): Hederella sp.
Brachiopoden (Armfüßer): Lingula sp. aff. brevis
Fuchs in Spriestersbach & Fuchs 1909, Stringocephalus
(Stringocephalus) sp., Retichonetes crenulatus (F.
Roemer 1844), Crurithyris inflata (Schnur 1853)
Muscheln: Ptychopteria (Actinopteria) reticulata
(Goldfuss), Leptodesma wupperana Herm. Schmidt 1909, Allorisma
sp. aff. westfalica (Spriestersbach 1917), Sphenotus
sp., Pteronites solenoides Herm. Schmidt 1909
Hyolithen: "Theca" sp.
Cephalopoden (Kopffüßer): "Gomphoceras"
sp.
Trilobiten: Dechenella burmeisteri Rud. Richter 1909
|
|
ca. 8 Meter |
dunkler, harter Riffkalkstein |
Stromatoporen:
Actinostroma
sp.
Rugose Korallen: Acanthophyllum sp., Disphyllum caespitosum (Goldfuss 1826)
Tabulate Korallen (Bödenkorallen): Thamnopora
sp.,
Favosites sp,
Coenites sp.
Crinoiden (Seelilien): Stielglieder
Brachiopoden
(Armfüßer):
Crurithyris inflata (Schnur 1853), Spinatrypa
aspera (Schlotheim 1813), Spiriferida (Reticulariacea indet.,
Spiriferacea indet.)
Muscheln:
Pterineidae
indet.
Ostracoden (Muschelkrebse):
Bairdiocypris sp.
|
|
ca. 20 Meter |
feinkörniger grauer bis brauner bankig-plattiger
Feinsandstein, teilweise mit laminierter Schichtung, nach oben
zunehmend kalkhaltig
|
Fauna aus Lesesteinen
Pflanzen: unbestimmbare Pflanzenreste
Ichnofossilien: Röhren- und Spreitenbauten
Brachiopoden (Armfüßer): Stringocephalus (Stringocephalus)
sp., Retichonetes crenulatus (F. Roemer 1844), Allorisma
sp. aff. westfalica (Spriestersbach 1917)
Trilobiten:
Dechenella burmeisteri Rud. Richter 1909
|
|
ca. 20 Meter |
grober Sandstein und graubrauner sandiger Tonstein mit
eingelagerten braunen mulmig verwitternden Siltsteinen
|
Fauna aus Lesesteinen
Pflanzen: unbestimmbare Pflanzenreste
Ichnofossilien: Chondrites sp.
Crinoiden (Seelilien): Stielglieder
Rugose Korallen: Heliophyllum sp.
Brachiopoden (Armfüßer): Spinatrypa aspera (Schlotheim
1813), Spinocyrtia (Carpinaria) ascendens (Spriestersbach
1935), ?Subcuspidella sp., Helaspis plexa (Wolfart
1956)
Gastropoden (Schnecken): Straparollus sp.
Muscheln: Ptychopteria (Actinopteria) reticulata
(Goldfuss)
Cephalopoden (Kopffüßer): "Orthoceras"
sp.
Trilobiten: Dechenella burmeisteri Rud. Richter 1909
|
|
Tab. 1: Schichtfolge am Zuckerberg.
|
Fossilführung
Landpflanzen
Da der Küstenverlauf des Nordkontinents während des Mitteldevon
nicht einheitlich war, bildeten sich, bedingt durch unterschiedliche
Wasserstände und ein periodisches Vordringen und Zurückweichen des
Meeres, vor der Küste Wattzonen, Überflutungsbereiche und
Flussdeltaräume mit teils marinen, teils limnisch-brackischen, teils
festländischen Bedingungen. Verbunden mit einem durch Äquatornähe
gleichmäßig warmen Klima boten diese Gebiete günstige Voraussetzungen
für die Entwicklung erster Landpflanzen, gewissermaßen für den
Schritt der Pflanzen vom Wasser aufs Land.
Die Landpflanzen des Mitteldevon waren einerseits sog. Psilophyten
(Nacktpflanzen); das sind blattlose Pflanzen, die auf eine zeitweilige
Überflutung ihres Lebensraumes noch nicht verzichten konnten. Weitere
Gruppen werden als Vorläufer der Farne, Schachtelhalme und
Bärlappgewächse gedeutet. Eine dritte Gruppe wird als Prospermatophyta
bezeichnet; diese fassen Formen zusammen, die sich zwar meist noch durch
Sporen fortgepflanzt haben, aber bereits durch ihren Holzbau zu den
Samenpflanzen überleiten. Sie besaßen kleine gegabelte Blättchen in
größeren Wedeln; teilweise waren sie baumförmig und verästelt. Die
Prospermatophyta werden als unmittelbare Ahnen der Samenpflanzen
angesehen.
Bei den in den Gesteinen am Zuckerberg auftretenden Pflanzenresten
handelt es sich z. T. um durch den Transport im bewegten Wasser
zerriebene Pflanzenteile (Häcksel), häufig in Lagen angereichert. Im
feinkörnigen Sandstein finden sich daneben gegabelte Achsen von
Psilophyten, z. B. Asteroxylon elberfeldense Kräusel &
Weyland 1926, aber auch Sprosse von Aneurophyton germanicum Kräusel
& Weyland 1923 und Protopteridium thomsonii Kräusel &
Weyland 1932, bei denen auch Zweigabschnitte mit kleinen spreitigen
Blättchen (Abb. 2a) und ansitzenden Sporangienträgern (Abb. 2b-c)
erhalten sind.
Während Asteroxylon elberfeldense
im Schwemmland des
nördlich gelegenen Flussdeltas gesiedelt haben dürfte, da es auf
Überflutungsperioden angewiesen war, stammen die anderen nachgewiesenen
Formen aus trockeneren Uferbereichen der Küstenebene.
|


|
Abb. 2: Sprosse
früher Landpflanzen (Vorläufer der Samenpflanzen).
a.-b. Aneurophyton
germanicum Kräusel & Weyland 1923;
Wedelabschnitte mit spreitigen
Blättchen,
a. Bildhöhe: 50 mm.
b. Bildbreite: 30 mm.
|
|


|
c.
Aneurophyton germanicum Kräusel & Weyland 1923; Wedelabschnitt
mit Sporangienträgern (rechts),
Bildbreite: 15 mm.
d.
Protopteridium thomsonii Kräusel & Weyland
1932; eingekrümmtes Wedelende mit Sporangienträgern,
Bildbreite: 15 mm. |
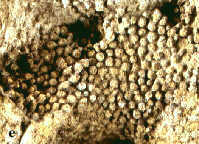
Korallen und Stromatoporen
Bei längerfristig ausbleibenden Sand- und Schlammschüttungen bildeten sich
zur Honsel-Zeit im flachen Schelfmeer örtlich Korallenriffe aus (siehe
oben). Die Riffbildung vollzog sich in sauerstoffreichem, gut durchlichtetem
Wasser bei einer Temperatur von 25°C. Ein bestimmter Salzgehalt des Wassers
war Voraussetzung, da Korallen im Süßwasser absterben und sich
Korallenriffe schon in weiterer Entfernung vor Flussmündungen auflösen.
Bei den in der Honsel-Formation vorkommenden
Riffen, die meist lokal begrenzt waren und eine Ausdehnung von mehreren
hundert Metern bis zu wenigen Kilometern hatten, handelt es sich um
Plateau-Riffe, d. h. geringmächtige Riff-Platten, die nur eine
vergleichsweise kurze Lebensdauer besaßen, da schon bald nach ihrer
Entstehung ihr Wachstum durch Sedimentschüttungen beendet wurde. Bildner
der Plateau-Riffe waren hauptsächlich Stromatoporen und Korallenkolonien;
die Riffoberfläche hatten ästige Korallen und Bödenkorallen besetzt.
Stromatoporen sind wahrscheinlich zu den
Schwämmen gehörende Lebewesen mit schichtiger Skelettstruktur (Abb. 3a).
Es kommen ausschließlich koloniale Formen mit massiven, knolligen,
kugeligen oder ästigen Stöcken vor. Während die Stromatoporen in der
Kreidezeit ausstarben, reichen die Korallen bis in die Gegenwart. Man
unterscheidet als wichtigste Ordnungen der im Devon vorkommenden Ordnungen
die Runzelkorallen (Rugosa) (Abb. 3b-c) und die Bödenkorallen (Tabulata)
(Abb. 3d-e). Rugose Korallen finden sich als Einzelkorallen und
Koloniebildner, während Bödenkorallen nur als Koloniekorallen auftreten.
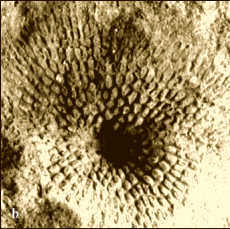
Moostierchen
Die Gruppe der Bryozoen (Moostierchen) lebt
auch heute noch. Es sind sessile, in Kolonien organisierte Strudler, deren
einzelne Individuen jedes für sich ein gelatinöses oder festes Gehäuse
aus Chitin bilden, z. T. zusätzlich mit Kalk. Erst durch ihre
Koloniebildung werden Bryozoen zu auffallenden Organismen. Sie sind am
Meersboden aufgewachsen oder bilden einen festen Überzug auf Steinen,
Pflanzenresten, Muschelschalen oder Krebspanzern. Im Mitteldevon sind u. a.
die Gattungen Fenestella und Hederella weit verbreitet und
kommen auch am Zuckerberg vor. Fenestella-Kolonien gleichen bei
vollständiger Erhaltung trichterförmigen Sieben (s. Abb. 4b), während Hederella
eine verästelte Wuchsform besaß (Abb. 4a). Moostierchen bevorzugten
tonreiche bis feinkörnig sandige Sedimente, kamen aber auch in Gesellschaft
von Korallenrasen oder Vorriff-Gebieten vor.
|

|
Abb. 4: Moostierchen.
a. Ästige
Bryozoe Hederella sp., auf eine Schale
aufgewachsen, Höhe: 10 mm.
b. Moostierchen-Kolonie Fenestella
sp.,
Durchmesser: 20 mm.
|
Brachiopoden
Brachiopoden (Armfüßer) besaßen im Erdaltertum große
Bedeutung, kommen aber, allerdings in geringer Zahl, auch heute noch in
verschiedenen Meeresbereichen vor.
Die bekannteste heute noch lebende Form ist
Lingula,
eine Gattung, deren Morphologie seit fünfhundert Millionen Jahren nahezu
unverändert geblieben ist. Linguliden gelten daher als „lebende
Fossilien". Sie gehören zu den inartikulaten (schlosslosen)
Brachiopoden, bei denen die beiden Klappen lediglich durch Muskeln zusammen
gehalten werden im Gegensatz zu den artikulaten (schlosstragenden)
Brachiopoden, zu denen die meisten Formen zählen. Auch am Zuckerberg konnte
die hornig-kalkige Schale einer Lingula (Abb. 5a) geborgen werden.
Oft werden Brachiopoden mit den Muscheln verwechselt, da beide Tiergruppen
über zwei Klappen verfügen. Während jedoch die Klappen der Muscheln meist
gleich sind und spiegelbildlich zueinander liegen, besitzen Armfüßer zwei
unterschiedliche Klappen: eine kleinere Armklappe und eine größere
Stielklappe. Aus letzterer tritt unterhalb des Wirbels durch eine Öffnung
ein fleischiger Stiel aus, mit dem sich das Tier am Boden festheftet. In der
Armklappe befinden sich zwei kalkige Armgerüste, mit denen es bei
geöffneter Schale Nahrung aufnimmt.
Aber auch der innere Körperbau beider Tiergruppen
unterscheidet sich stark, so dass Muscheln und Brachiopoden nicht einmal
entfernt miteinander verwandt sind: Muscheln zählen wie die Schnecken zu
den Weichtieren (Mollusken), Brachiopoden dagegen bilden einen eigenen Stamm
im Tierreich.
Auch bei den Funden vom Zuckerberg kommen Brachiopoden
arten- und individuenreich vor (siehe Tabelle 1). Zahlenmäßig überwiegt
die Gattung Retichonetes (Abb. 5b), die im Feinsandstein z. T.
Schilllagen bildet. Retichonetes und Verwandte gehören zum
ökologischen Grundtyp der freiliegenden Brachiopoden, das heißt: Bei
diesen Formen erfolgte keine Fixierung am Meeresboden mittels eines Stiels,
vielmehr erhielten Chonetiden ihre Stabilität durch relativ lange Stacheln
am Schlossrand, mit denen sie dem Sediment auflagen. Zusätzlich geschützt
und damit auch stabilisiert waren sie durch die halb eingegrabene Lage ihres
Gehäuses im Sediment.
Einige Brachiopoden-Gruppen stellen wichtige Leitfossilien
im Mitteldevon dar, die bei genauer Bestimmung Rückschlüsse auf das genaue
Alter zulassen. Einige kommen auch am Zuckerberg vor; allerdings ist die
vorliegende Steinkernerhaltung meist nicht ausreichend, um
feinstratigraphische Aussagen treffen zu können (siehe oben: Kap. „Die
Schichtfolge" und Anm.2).
Häufig in der Honsel-Formation vorkommende Brachiopoden gehören zur
Ordnung Spiriferida, deren Vertreter ein spiralig aufgerolltes Armgerüst
besitzen. Eine charakteristische Form aus dieser Ordnung ist Spinocyrtia
(Carpinaria) ascendens (Abb. 5c). Ebenfalls zur Ordnung
Spiriferida (Unterordnung Atrypidina) gehören zahlreiche mitteldevonische
Gattungen. Am Zuckerberg treten insbesondere Formen aus der Spinatrypa
aspera-Gruppe auf, die durch ihre berippte, schuppig gefaltete und
bestachelte Schale auffallen (Abb. 5f).
|
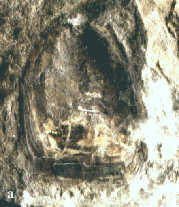 
|
Abb. 5: Brachiopoden.
a. Lingula sp. aff. brevis Fuchs in
Spriestersbach & Fuchs 1909,
Länge: 16 mm.
b. Retichonetes crenulatus (F. Roemer 1844),
Schilllage, Bildbreite: 45 mm.
|
|


 
|
c. Spinocyrtia (Carpinaria) ascendens (Spriestersbach
1935), Breite: 24 mm.
d. Stringocephalus (Stringocephalus) sp. (Steinkern der
Armklappe), Breite 25 mm.
e. Athyris sp., Breite: 16 mm.
f. Spinatrypa aspera (Schlotheim
1813), Breite: 25 mm.
|
Ringelwürmer
Reste von Ringelwürmern (Anneliden), zu denen auch der
Regenwurm zählt, sind recht selten. Gelegentlich findet man auf Schalen von
Meerestieren nur wenige Millimeter große planspiral aufgewundene Röhren,
die einem schneckenartigen Gehäuse gleichen. Die Bildungen stammen von
Ringelwürmern der Gattung Spirorbis (Abb. 6). Diese Gattung ist seit
dem Ordovizium nachgewiesen und existiert heute noch. Häufiger sind auch Schalen von Brachiopoden oder Muscheln von
zylindrischen Bohrgängen durchzogen, die sich insbesondere als Abdruck auf
Steinkernen finden und ebenfalls auf Ringelwürmer zurückgeführt werden.
|

|
Abb. 6: Eingerollte Wurmgehäuse der Gattung Spirorbis,
aufgewachsen auf einer Muschelschale, Durchmesser des
größten Exemplars: 5 mm.
|
Schnecken
Schnecken (Gastropoden) existieren zwar seit dem frühen
Erdaltertum und treten bereits im Kambrium auf. Sie bleiben jedoch zunächst
stammesgeschichtlich, wie die Muscheln auch, von geringer Bedeutung. Erst
mit dem Erdmittelalter werden die Schnecken häufiger, auch die
Formenvielfalt und Mannigfaltigkeit der Verzierungen nimmt zu. In der
Tertiärzeit erleben sie das Maximum ihrer Entwicklung und überflügeln die
Muscheln an Häufigkeit.
Im sandig-tonigen Mitteldevon gehören Schnecken zu den
seltenen Funden; und so konnten am Zuckerberg auch nur zwei Formen eindeutig
nachgewiesen werden: Die Gattung Straparollus (Abb. 7a) mit
planspiralem Gehäuse, bei dem die Mittellinie aller Windungen in einer
Ebene liegt, und die Gattung Loxonema (Abb. 7b) mit trochispiral
gewundenem Gehäuse. Bei dieser Form legen sich die Windungen von der Spitze
an spiralartig aneinander und werden von Umgang zu Umgang stetig breiter.
|


|
Abb. 7: Steinkerne
von Schnecken.
a. Straparollus sp., Durchmesser des Gehäuses:
8 mm.
b. Loxonema sp., Höhe: 7 mm.
|
Muscheln
Wie die Schnecken so haben auch die Muscheln
in den Meeren des Erdaltertums im Vergleich zu den Meeren der Neuzeit eine
nur untergeordnete Bedeutung. Dennoch sind sie auch in früher Zeit bereits
formenreich vertreten. Bei aller Formenfülle stellen sie jedoch im
Vergleich zu anderen Fossilgruppen nur wenige Leitfossilien. Dies liegt u.a.
auch an ihrer Faziesabhängigkeit. Aber gerade diese für die
Biostratigraphie ungünstige Eigenschaft macht die Muscheln besonders
wertvoll für die Ausdeutung des Lebensraumes. Zum Beispiel unter lagunären
Bedingungen, in Nähe des Festlandes und in Deltabereichen kamen sie
häufiger vor, so auch in den Flachwasserablagerungen der Honsel-Formation.
Am Zuckerberg findet sich im Feinsandstein
die Art Leptodesma wupperana (Abb. 8c) teilweise sogar in
Anhäufungen, während die großen Exemplare von Ptychopteria (Actinopteria)
reticulata (Abb. 8a) und Allorisma sp. aff. westfalica
(Abb.8b) eher selten und vereinzelt zu finden sind.
|




|
|
|
Abb. 8: Muscheln.
a.
Ptychopteria (Actinopteria) reticulata (Goldfuss),
Bildbreite: 65 mm.
b. Allorisma sp. aff. westfalica (Spriestersbach 1917),
Bildbreite: 35 mm.
c. Leptodesma wupperana
Herm. Schmidt 1909.
d. Sphenotus sp. (beide Klappen erhalten),
Bildbreite: 25 mm.
|
Cephalopoden
Eine Tiergruppe, der seit dem Devon besondere
Bedeutung zukommt, sind die Cephalopoden (Kopffüßer), die nicht nur die
höchstentwickelten Weichtiere (Mollusken) sind, sondern auch die größten
bekannten Tierformen unter den Wirbellosen.
Der einzige heute noch lebende Kopffüßer
mit Außengehäuse ist der in südostasiatisch/australischen Meeren
vorkommende Nautilus; andere rezente Formen wie Kraken und Kalmare
besitzen kein Außengehäuse, sondern lediglich eine hornartige Platte unter
dem Mantel. Die bekanntesten fossilen Cephalopoden sind die aus der Jura-
und Kreidezeit stammenden Ammoniten.
Die häufigsten Cephalopoden im Mitteldevon
sind gestreckte oder leicht gekrümmte Nautiliden, am bekanntesten die
schlank zylindrischen Orthoceren.
Bei den am Zuckerberg entdeckten Nautiliden
handelt es sich um die Gattungen „Orthoceras" (gestreckt), Rutoceras
(leicht gekrümmt) und „Gomphoceras" (birnenförmig), die alle
aber nur fragmentarisch erhalten sind.
|

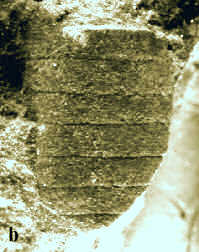
|
Abb. 9: Bruchstücke
von Nautiliden.
a. Rutoceras sp., Länge: 20 mm.
b.
„Orthoceras" sp., Länge: 10 mm.
|
Hyolithen
Bei den Hyolithen handelt es sich um
Organismen mit spitz-konischem Gehäuse, die nur im Kambrium und Ordovizium
häufiger, sonst aber recht selten sind und nur mit etwa 20 Gattungen
nachgewiesen wurden. Sie traten im Kambrium auf und starben im Perm wieder
aus. Da man sehr wenig über ihren inneren Körperbau weiß und ihre
Lebensweise nicht kennt, ist ihre systematische Stellung im Tierreich nicht
eindeutig geklärt. Die Zugehörigkeit zu den Mollusken lässt sich von der
Schalenstruktur und von der gelegentlich beobachteten Kammerung her aber
annehmen.
Die im Steinbruch Zuckerberg geborgenen
Exemplare haben große Ähnlichkeit mit der von Spriestersbach (1942: 147
und Taf. 1) aus dem Unteren Mitteldevon beschriebenen und abgebildeten
Gattung Theca. Es handelt sich um Steinkerne, die völlig glatt sind
und keine Skulptur oder Anwachsstreifen erkennen lassen.


|
Abb. 10: Hyolithen.
a.-b. Unvollständig erhaltene Exemplare
(Steinkerne) der Gattung "Theca", Höhe: 14 mm.
|
Tentaculiten
Die Tentaculiten sind eine bereits im
Oberdevon weitgehend erloschene Tiergruppe. Ihre systematische Stellung
ließ sich erst vor einigen Jahren einigermaßen absichern, nachdem man
erhaltene Weichkörperreste mit Hilfe von Röntgenstrahlen untersuchen
konnte. Danach werden sie als eigene Klasse zu den Weichtieren (Mollusken)
gewertet, die insbesondere den Kopffüßern (Cephalopoden) nahe steht.
Mit ihren millimetergroßen
spitzkegelförmigen Gehäusen schwammen sie in teilweise großen Schwärmen
im Meer. Häufig findet man sie durch die Strömungsrichtung eingeregelt im
Gestein.
In den Schichten vom Zuckerberg konnte
allerdings nur sehr wenige Exemplare entdeckt werden.
Seelilien
Die heute noch vorkommenden Seelilien (Crinoiden)
gehören zusammen mit Seeigeln, Seesternen und anderen zum formenreichen
Stamm der Stachelhäuter. Seelilien besitzen einen Stiel, mit dem sie im
Boden verankert sind. Darauf befindet sich ein Kelch mit Armen, mit denen
Nahrung eingestrudelt wird. Kelch und Arme bilden die „Krone". Nach
dem Absterben des Tieres zerfällt meist dieses Skelett, weshalb komplette
Kronen oder Kelche fossil nur selten entdeckt werden können. Meist findet
man isolierte Glieder, die den langen Stiel aufbauten (Abb. 11).
In flachen Meeren bildeten Seelilien
ausgedehnte „Wälder". Seelilien lassen sich aber in vielen
Ablagerungen des devonischen Schelfmeeres auffinden, da insbesondere die
Stielglieder sehr leicht und weit verdriftet wurden. So kommen auch am
Zuckerberg Stielglieder von kleinwüchsigen Exemplaren in zum Teil
zusammengeschwemmten Lagen im groben Sandstein vor.
|

|
|
Abb. 11: Seelilien-Stielglieder;
Bildbreite: 30 mm. |
Ostracoden
Ostracoden (Muschelkrebse) werden meist als
eine eigene Tierklasse innerhalb der Krebstiere (Crustacea) angesehen. Sie
besitzen eine zweiklappige, kalkige und meist asymmetrische Schale. Die
Tiere sind sehr klein, nur etwa 0,5 bis 5,0 mm lang. Einige „Riesenformen"
aus dem Silur bringen es aber auf bis zu 3 cm Länge. Auf den ersten,
oberflächlichen Blick erinnert die Schale an eine winzige Muschel, was den
Tieren ihren deutschen Namen eingebracht hat. Dies gilt insbesondere für
glattschalige Formen; oftmals tragen Ostrakoden-Klappen aber auch
auffällige und kennzeichnende Verzierungen.
Die Ostracoden wurzeln im Kambrium und
reichen bis in die Gegenwart, in der sie das Maximum ihrer Formenvielfalt
haben. Ursprünglich kamen sie nur im Meer vor, seit Ende des Devon aber
auch in Süßwasserbereichen. Sie leben sowohl freischwimmend als auch
bodenbewohnend.
In den Schichten am Zuckerberg konnten nur
wenige Exemplare gefunden werden. Während die Stücke aus dem groben
Sandstein wegen schlechter Erhaltung unbestimmbar sind, handelt es sich
einem Exemplar aus dem Riffkalk um die Gattung Bairdiocypris. Diese
kommt in den Schichten des Oberen Mitteldevon nur vereinzelt vor und gehört
zu den glattschaligen Formen, die typisch sind für den lagunären
Ablagerungsraum der Honsel-Formation.
|
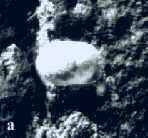

|
Abb. 12: Ostracode Bairdiocypris
sp.,
Länge: 2 mm.
a. Foto.
b. Seitenansicht der rechten Klappe (Zeichnung).
|
Trilobiten
Wie die Ostracoden, gehören die im Kambrium
auftretenden und am Ende des Erdaltertums (Perm) aussterbenden Trilobiten
zum Stamm der Gliederfüßer (Arthropoden). Mit den Krebsen aber sind sie
nicht näher verwandt, wenn auch als deutsche Bezeichnung gelegentlich der
Name „Dreilappkrebs" verwendet wird. Das Wort „Trilobit"
bedeutet soviel wie „Dreilapper". Namengebend ist nämlich die
auffällig dreiteilige Längs- und Quergliederung des Rückenpanzers: Von
vorn nach hinten besteht der Panzer aus Kopfschild (Cephalon), Rumpf
(Thorax) und Schwanzschild (Pygidium). In der Querrichtung lassen sich die
Mittelachse oder Spindel (Rhachis) und die Seitenfelder (Pleuralfelder)
unterscheiden. Der Kopfschild ist durch die beiden Gesichtsnähte (Facial-Suturen)
nochmals in drei Felder unterteilt: An den Mittelkopf (Cranidium) mit
Glabella und rechter wie linker Festwange schließen sich links und rechts
die Freiwangen mit Auge und gegebenenfalls Wangenstacheln an. Hinzu kommt von der Kopf-Unterseite als
erhaltungsfähiges Hartteil der Mundschild (Hypostom).
Da der Panzer nicht mit dem Weichkörper des
Tieres mitwachsen konnte, musste es sich mehrfach (bis zu 30 mal) in seinem
Leben häuten. Solche Häutungsreste, meist verdriftete Panzerteile, sind
daher viel häufiger zu entdecken als vollständige Panzer gestorbener
Tiere.
So findet man am Zuckerberg von der einzigen
vorkommenden Art Dechenella burmeisteri auch nur Panzerteile: meist
Pygidien (Abb. 13c), seltener Cranidien (Abb. 13d) und Freiwangen (Abb.
13e). Manchmal findet man auf einer Platte mehrere Panzerteile: So liegt ein
Stück vor mit Cranidium, Freiwange und Hypostom. Das Exemplar (Abb. 13a),
bestehend aus Pygidium und Thorax im Zusammenhang (Thoracopygidium) gehört
bei der Gattung Dechenella durchaus zu den seltenen Funden.
Trilobiten haben im Laufe ihrer Entwicklung
zahlreiche Leitfossilien hervorgebracht. Auch Dechenella burmeisteri
gehört dazu; die Art kommt zwar gelegentlich an wenigen Fundorten in den
obersten Schichten der Unterhonsel-Formation vor, sonst aber zahlreich nur
in der Oberhonsel-Formation. Am Zuckerberg gehören Panzerteile von Dechenella
burmeisteri zu den häufigen Funden.
|

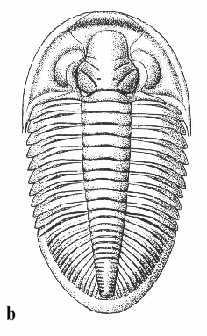
|
Abb. 13: Trilobiten.
a. Thorax und Pygidium eines ausgewachsenen Exemplars von Dechenella
burmeisteri Rud. Richter 1909;
Länge: 30 mm.
b.
Rekonstruktionszeichnung eines kompletten Dechenellen-Panzers.
|
|



|
c.-e. Panzerteile von Dechenella
burmeisteri Rud. Richter 1909 (Jugendexemlare).
c. Pygidium, Breite:10 mm.
d. Freiwange mit
Auge und Wangenstachel,
Bildhöhe: 20 mm.
e. Cranidium (Mittelkopf), Bildhöhe:10 mm.
|
Ichnofossilien
Bei einigen der vorkommenden Fossilien handelt es sich nicht um Reste der
Tiere selbst, sondern um Überbleibsel ihrer Lebenstätigkeit (=
Ichnofossilien), die bestimmte Organismen beim Graben im Sediment, auf
Schalen anderer Tiere, in Pflanzenresten, bei der Nahrungsaufnahme oder als
Kot- oder Eiablage hinterlassen haben. Meist sind die Erzeuger solcher
Spuren unbekannt, werden auch möglicherweise nie gefunden werden, da sie
keine erhaltungsfähigen Hartteile besitzen.
Hauptsächlich kommen im groben Sandstein am
Zuckerberg verzweigte Gänge (Fressbauten) der Gattung Chondrites vor;
möglicherweise wurden diese Spuren von wurmartigen Lebewesen verursacht,
die als Sedimentfresser das Sediment nach Nahrung durchpflügten. Ihre
Tunnelsysteme durchlaufen das Sediment zunächst quer zur Schichtung; erst
die Gangenden liegen dann parallel zur Schichtung. Der Durchmesser der
Röhren beträgt meist 0,5 mm, vereinzelt aber auch 5 mm (Abb. 14).
|

|

|
Abb. 14: Chondrites sp.
(Grabgänge).
a. Bildbreite: 60 mm.
b. Bildbreite: 80 mm.
|
Problematica
Als Problematica werden solche Fossilien
bezeichnet, die über keine Merkmale verfügen, nach denen sie einer
bestehenden Tier- oder Pflanzengruppe eindeutig zuzuordnen sind. So kommen
auch am Zuckerberg Abdrücke mit schalenartigem Aussehen vor, die zwar
Muschel- oder Brachiopodenklappen ähnlich sehen, doch andere
Symmetrieverhältnisse besitzen. Hierbei könnte es sich um
Phyllocariden-Panzer handeln (Abb. 15). Diese auch als Blattkrebse
bezeichnete Gruppe der Höheren Krebse (Malacostraca) besitzt einen
zweiklappigen Carapax, der den Körper seitlich schützt.
Phyllocariden-Funde bestehen meist aus solchen isolierten Klappen, seltener
aus Teilen des Körpers (Thorax-Segmente oder Teile des Abdomens);
allerdings fehlen bei den am Zuckerberg entdeckten Stücken wichtige
Merkmale, die eine klare Zuordnung zu dieser Gruppe ermöglichen würde.
Erst weitere besser erhaltene Funde könnten zur Klärung des Problems
beitragen.
|

|
Abb. 15: ?Phyllocariden-Panzer indet., Breite: 20 mm.
|
Problematisch sind auch Strukturen in einem
Holzrest (Abb. 16), die vorläufig als Fraßspuren gedeutet werden. Da das
Stück keine 2 cm groß ist, sind die Spuren sehr filigran. Sie sind sowohl
in der Negativ- als auch in der Positivplatte negativ erhalten.
Allerdings wäre auch denkbar, dass es sich
um Bryozoen-Aufwuchs handelt, insbesondere aufgrund der kolonieartigen
Organisation. Die härteren Strukturen der Bryozoen-Kolonie (die dann
später weggelöst wurden) könnten sich infolge des bei der Diagenese
entstehenden Druckes von außen negativ in die weiche Pflanzensubstanz
durchgeprägt haben.
|

|
Abb. 16: ?Spuren in einem Holzrest,
Bildhöhe 14 mm.
|
|
Abbildungen
Abgebildete Fossilien: Sammlungen L. Koch (29), U. Lemke (5), Arbeitskreis
Kluterthöhle (1)
Fotos: L. Koch (43), U. Lemke (2)
|
|
Abkürzungen
aff. (lat. affinis) angrenzend; Zeichen der offenen Namengebung: „aus der
Verwandtschaft von".
|
|
indet. (lat.
indeterminabel) unbestimmt, unbestimmbar; hinter einem Ordnungs- oder
Familienname: nicht bis zur Gattung und Art zu bestimmen.
|
|
sp. (lat. species) Art; nach einem Gattungsnamen: die Art ist nicht zu
bestimmen.
|
|
? Ein Fragezeichen vor einem Begriff zeigt an, dass gewisse Zweifel an der
Richtigkeit bestehen.
|
|
"....." Ein Gattungsname in Anführungszeichen gibt an, dass es
sich um eine ältere Sammelgattung handelt, eine Zuordnung zu neu
aufgestellten Gattungen aber nicht möglich ist.
|
|
Anmerkungen
1 Die projektierte Bundesstraße 7e war als
Entlastungsstraße zur B7 zwischen Schwelm und Ennepetal geplant, kam aber
nicht zur Ausführung. Das Erdbaulaboratorium Ahlenberg, Herdecke, führte
im Auftrag des Landesbetriebs Straßenbau NRW, Bochum, Anfang 1996 am Fuße
des Zuckerberges mehrere Kernbohrungen zur Analyse des Untergrundes bis zu
einer Teufe von ca. 20 m durch.
|
|
2 Messtischblatt 1:25.000 Barmen; Koordinaten: r2592625,
h5685275.
|
|
3 Die vorkommenden Exemplare von Spinatrypa lassen sich
lediglich in der Sammelart aspera (Schlotheim 1813) zusammenfassen,
während in der Kluterthöhle die Art Spinatrypa orthoclina Copper
1967 gefunden wurde und die Einstufung der Schichten in die
Oberhonsel-Formation belegt (vgl. Koch 1992: 29: Abb. u.l.).
|
|
4 Conodonten sind zahnartige, lamellar aufgebaute Mikrofossilien
aus Calciumphosphat. Sie erreichen Größen bis zu 3 mm und haben ein hohes
spezifisches Gewicht (2,84). Sie gehören als bevorzugt erhaltungsfähige
Hartsubstanzen, als Skelett-Elemente zu einer ausgestorbenen ursprünglichen
marinen Wirbeltiergruppe.
Conodonten finden sich in Sedimentgesteinen vom jüngsten Präkambrium bis
in die Trias. Sie gehören zu den wichtigsten Zeitmarken, mit denen
Sedimente diesen Alters in der Praxis datiert werden. Aus Kalksteinen des
Devon können Conodonten mittels Essigsäure herausgelöst werden. An den
isolierten Conodonten werden Bestimmungen und, daraus resultierend, genaue
Altersdatierungen durchgeführt.
|
|
5 Kernbohrung des Erdbaulaboratoriums Ahlenberg (s. Anm. 1); r2592527, h5685433.
|
|
Literatur
Çinar, C. (1978): Marine
Flachwasserfazies in den Honseler Schichten (Givet-Stufe, Lüdenscheider Mulde,
Rechtsrheinisches Schiefergebirge). – Göttinger Arbeiten für Geologie
und Paläontologie, 20: 1-77, Abb.1- 28, Profile 1-4, Taf. 1-11; Göttingen.
|
|
Fuchs, A. & Paeckelmann, W. (1979): Geologische Karte von
Nordrhein-Westfalen 1: 25000, Erläuterungen zu Blatt 4709 Wuppertal-Barmen, 2.
Aufl.: I-VII und 1-96, 7 Abb., 1 Tab., 1 Taf.; Krefeld. |
|
Koch, L. (1992): 380 Millionen Jahre Erdgeschichte. Der Klutertberg und seine
geologische Entwicklung. –
in: Koch, L. (Hg.): Das Klutert-Buch: 10-35, 36 Abb.; Hagen (v. d.
Linnepe). |
|
Langenstrassen, F.
(1982): Sedimentologische und biofazielle Untersuchungen an Proben aus der
Bohrung Schwarzbachtal 1 (Rheinisches Schiefergebirge, Velberter Sattel). –
Senckenbergiana lethaea, 63 (1/4): 315-333, Abb. 1-2, Taf. 1-2; Frankfurt am
Main.
|
|
May, A. (1986): Biostratigraphische Untersuchungen im Mittel-Devon des
Nordwest-Sauerlandes (Rheinisches
Schiefergebirge). – Dortmunder Beiträge zur Landeskunde,
naturwissenschaftliche Mitteilungen, 20: 23-55, Abb. 1-2, Tab. 1-2;
Dortmund. |
|
Spriestersbach, J. (1942): Lenneschiefer
(Stratigraphie, Fazies und Fauna). –
Abhandlungen des Reichsamts für Bodenforschung, Neue Folge, 203:
218 S., 19 Abb., 11 Taf.; Berlin.
|
|
Weddige,
K. [Hg.] (1996): Devon-Korrelationstabelle. – Senckenbergiana lethaea, 76 (1/2): 267-286, Abb. 1-7, 43
Tab.-Spalten; Frankfurt am Main.
>>
zur
Hauptseite |
|